Correlations
| North Pacific | AKSR-NASR | Anadyr-Koryak | Verkhoyansk-Chukotka | Okhotsk-Chukotka | N. Alaska | Yukon | Chignik |
|---|---|---|---|---|---|---|---|
| Phytostratigraphy | Images of fossils characteristic of the Type Taphoflora | ||||||
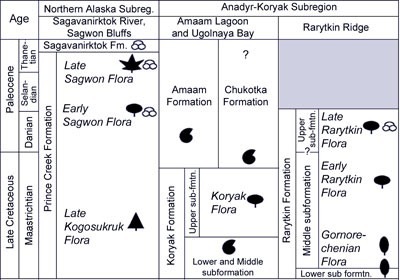
The Koryak Phase of Floral EvolutionBudantsev (1983), who was first to distinguish this phase, attributed to it a series of the Coniacian–Danian floras known in northern and northeastern Asia and North America and named the phase after the Koryak taphoflora of the Amaam Lagoon area. Subsequently, Herman (1988, 1993, 1999a) considerably narrowed the stratigraphic and geographic ranges of the phase, though retained the given name following the nomenclatural principle, adopted in systematics, of priority. Golovneva (1994a) argued that a flora from the upper half of the Rarytkin Formation characterizes better the age range of the phase and suggested a retrospective renaming of the phase. This Rarytkin flora probably is more representative but changes in nomenclature cannot be motivated by new data on the phase range. In biology, for instance, nomenclatural changes do not occur following new information about the name-bearing type species/holotype. Changes are only invoked if earlier valid type material is found or as a result of new combinations of material. Following Golovneva, the same argument in favor of using the name "Rarytkin" for the phase has been repeated by Pokhialainen (1994), (pp. 17–18): "… flora of the Rarytkin Formation corresponds in greatest extent to the phase under consideration… In this connection, the Santonian–Maastrichtian plant assemblages of coastal plains … can be termed the Rarytkin stratoflora…". However, following nomenclature principles of typification and priority, the original term suggested by Budantsev (1983), who named the phase after the Koryak taphoflora of the Amaam Lagoon area, is used here. In addition to its type flora (Golovneva and Herman, 1992; Herman, 2011a; Moiseeva, in press), the Koryak phase includes the Rarytkin taphofloras known in the northern and southwestern areas of the Rarytkin Ridge, and that on the western flank of the Pekul'nei Ridge (Golovneva, 1994a). The last one consists of seven species only, six of which are characteristic of the Gornorechenian phase also, and can be only tentatively attributed to the Koryak phase. Golovneva (1994a) also included in the Koryak phase the floral assemblage of the Kholminskaya succession in the Khatyrka River basin, where that assemblage is again too impoverished and featureless to be confidently attributed to any phase. The same can be said about taphofloras from the Rarytkin Formation of the Pekul'nei Ridge eastern slope, the topmost interval of the Koryak Formation in the Ugol'naya Bay site, and the upper part of the Belorechenskaya Formation in the southwestern area of the Rarytkin Ridge. The boundary between the Gornorechenian and Koryak phases can be established based on the succession of taphofloras in the middle Rarytkin Subformation exposed on the northern part of the Rarytkin Ridge. This stratigraphic interval is, however, lacking in marine deposits with stratigraphically important mollusk remains, and despite the observable succession of taphofloras, the position of the boundary that separates them is problematic. This is because the taphofloras of the Gornorechenian and Koryak phases are separated by a 200 m-thick interval devoid of plant remains. On the other hand, the Koryak taphoflora of the Amaam Lagoon area is correlative with the Rarytkin floral assemblage found at a higher level, while the Kakanaut taphoflora of the Pekyl'neiskoe Lake area is comparable in composition with plant remains of the lower Gornorechenian phase. Based on relations with marine deposits, both taphofloras correspond in age to the Maastrichtian and the boundary between the Gornorechenian and Koryak phases is placed in, approximately, the middle of the Maastrichtian. The upper boundary of the Koryak phase is difficult to place because it cannot be substantiated by independent (non-paleobotanical) methods. It is within the interval spanning the end of the Maastrichtian to the end of the Danian. In the opinion of Golovneva (1994a), this phase (Rarytkin stratoflora in her terminology) spans the terminal Maastrichtian–Danian interval because the taphoflora of the upper Rarytkin Subformation is of Danian age based on its taxonomic composition. The Koryak phase most likely lasted 5 - 8 m.y. as far as can be determined on current evidence. Plant taxa characteristic of the flora are listed in the panel to the right. Taphofloras of the Koryak phase are dominated by Metasequoia, Corylites beringianus, Trochodendroides (five species), and in some localities by Glyptostrobus, Mesocyparis, and Ettingshausenia raynoldsii. The other characteristic, though less abundant, taxa of the Koryak flora are Equisetum arcticum, Onoclea hesperia, Elatocladus talensis, Araucarites conophyllus, Sequoia amaamensis, Haemanthophyllum cordatum, Celastrinites septentrionalis, Cissites volobuevae, and Rarytkinia terechovae. The abundance of large-leaved angiosperms (Ettingshausenia, Arthollia, Corylites, "Pterospermites") is higher than in the Gornorechenian phase. Conifers, mostly Cenozoic genera, are important components often representing the dominant subgroup in addition to angiosperms. Horsetails and ferns are not numerous, and Ginkgo remains are extremely rare; cycadophytes are absent. In general, the flora of the Koryak phase is already close in taxonomic composition to the Danian–Paleocene but not the Late Cretaceous floras of Northeastern Asia and North America (Vakhrameev and Akhmet'ev, 1977; Budantsev, 1983; Golovneva, 1994a; Golovneva and Herman, 1992; Moiseeva, in press). It is likely that the late Maastrichtian flora of the AKSR was a source, if not the origin, of the Early Tertiary deciduous floras of the Arctic and adjacent regions (Boulter et al., 1988; Herman and Spicer, 1995, 1997a). |
|
||
|